Степени СВОБОДЫ Ума. Урок ПОНИМАНИЯ 3.
Просмотров: 1828
Вы пока не авторизованы на сайте.
Войдите на сайт или зарегистрируйтесь, чтобы добавлять свои материалы на сайт.
Открытие, мимо которого прошло не одно поколение генетиков XX столетия «В большинстве случаев одного простого опыта достаточно, чтобы установить самое главное» / Луи Пастер
Чуть более двух лет назад в статье под скучным названием «Необычное поведение полового тельца (ПТ) в сперматогенезе у мышей, подвергшихся мутагенному воздействию», были представлены результаты, свидетельствующие о том (Захидов и др., 2020), что нами впервые в мире установлен новый тип крупномасштабной хромосомной аномалии, четко выявляемой микроскопически на стадии профазы I мейоза. Это – индивидуализация полового бивалента XY (или полового тельца), его резкий отрыв от основного хромосомного материала развивающийся мужской половой клетки.
Причина такого катастрофического пространственного перемещения гетерохромосомной пары XY из ядра в цитоплазматическую среду клетки не совсем ясна. Предположительно она может быть связана с индуцированными нарушениями внутриядерных физико-химических взаимодействий, в норме играющих ведущую роль в обеспечении фундаментальной устойчивости и упорядоченности хромосом в клетках, движущихся по сложному мейотическому циклу (Захидов и др., 2020).
Как бы там ни было, но разрыв между основным ядром и половым бивалентом XY, представляющим собой консолидированную замкнутую структуру, не приводил к его выбраковки и элиминации из клетки. Напротив, структура XY, потеряв глубинную общность с основным генетическим аппаратом, проявила поразительную устойчивость и топологическую цельность, она не устранялась из генеративного аппарата, и не оставалась чужой даже после делений мейоза, легко принималась постмейотическими клетками – сперматидами и сперматозоидами.
Объяснение длительному автономному существованию гетерохромосомной пары XY при химической индукции мутаций следует искать в том, что в сложных протоплазматических средах целые отдельные хромосомы или их крупные фрагменты сохраняют известное время свое генетическое состояние (Рапопорт, 1991).
Важно заметить, что крупномасштабная мутация в структуре хромосомного материала, хотя и изменяла общий хромосомный баланс, скорее всего, не приводила к нарушению процесса формирования жизнеспособных, морфологически нормальных, но «бесполых» сперматозоидов, т.е. сперматозоидов, лишенных половых хромосом – X или Y (Захидов и др., 2020).
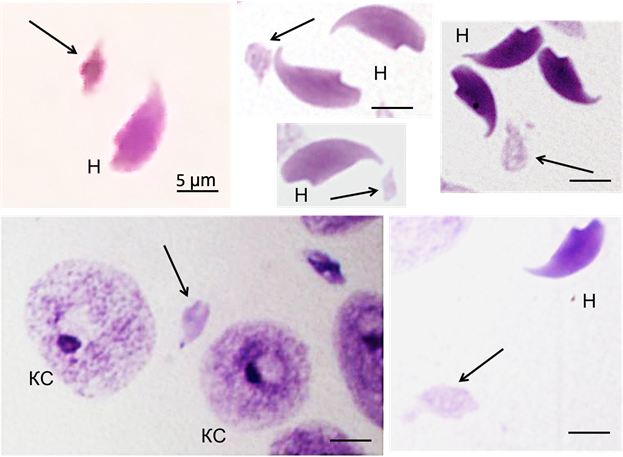
В настоящем сообщении нами представлены новые наблюдения, демонстрирующие, что в семенниках мутагенизированных мышей, половой бивалент XY, выведенный за пределы естественного ядерного окружения и потерявший всякую возможность вновь объединяться с основным ядром в единое тело, обнаружил способность трансформироваться в структуру, напоминающую ядро морфологически нормального сперматозоида (рис. 1).
Рис. 1. Спермии мутагенизированных мышей-гибридов F1 CBA × C57BL/6. Обозначения: Н – нормальные ядра, стрелками указаны аномальные ядра с генетической структурой XY; КС – ядра вспомогательных клеток Сертоли. Окраска по Фельгену.
Вопрос о том, могут ли эти необычные миниспермии, лишенные аутосом и состоящие исключительно только из двух половых хромосом X и Y, транспортироваться в женские половые органы, и далее внедряться в яйцеклетки, пока остается открытым.
Однако если теоретически допустить, что миниспермии с генетической структурой 0 + XY в состоянии по отдельности или совместно с нормальными гаплоидными спермиями 19 + X или 19 + Y оплодотворять нормальное яйцо 19 + X, то тогда возможны следующие варианты хромосомной несбалансированности оплодотворенной яйцеклетки: 19 + XXY и 38 +XXXY или 38 + XXYY (нормальный диплоидный кариотип мыши состоит из 40 хромосом, из них 38 аутосом и 2 половые хромосомы, соответственно XX – у самки и XY – у самца).
С другой стороны, если предположить, что нормальная гаплоидная яйцеклетка 19 +X будет оплодотворена «бесполым» сперматозоидом 19 + 0, то впоследствии мы можем получить организм с кариотипом 38 + X0.
Как известно, в большинстве случаев возникновение количественного дисбаланса хромосом после слияния мужской и женской половой клетки может иметь следствием ненормальное развитие зародыша, спонтанные аборты или появление на свет генетически нездорового потомства.
В этой связи нельзя не сказать о том, что, например, с точки зрения частной генетики человека, увеличение числа людей, отягощенных вредными наследственными признаками, в том числе под влиянием радиации (высокоэнергетических квантов и элементарных частиц) или молекулярных мутагенов, представляет серьезную угрозу существованию человеческого рода.
Итак, как явствует из наших суммарных наблюдений, нарушение топологической композиции генома на уровне полового бивалента XY может приводить к формированию двух типов аберрантных гамет: миниспермиев с уникальным хромосомным комплексом 0 + XY и «бесполых» спермиев с генетической структурой 19 + 0.
В целом же вся совокупность полученных нами экспериментальных данных о поведении и судьбе гетерохромосомной пары XY (полового тельца) в условиях потери ею оптимального состояния, открывает новую страницу в генетической науке, объектом изучения которой являются, не в последнюю очередь, половые клетки, обеспечивающие наследственную преемственность в ряду поколений.
Также результаты наших исследований подтвердили один из постулатов, сформулированных в свое время выдающимся советским генетиком М.Е. Лобашёвым: «Как часто бывает в науке, классическое открытие делается на элементарном, на первый взгляд, явлении и при использовании простой методики».
Литература.
Захидов С. Т., Муджири Н. М., Макарова И. В., Андреева Л. Е. Необычное поведение полового тельца (ПТ) в сперматогенезе у мышей, подвергшихся мутагенному воздействию» // Известия РАН, Сер. Биол., 2020, № 6, с. 581–585.
Рапопорт И. А. Генетическая дискретность и механизм мутаций // В кн.: Химический мутагенез и проблемы селекции. – М.: Наука, 1991.– С. 3–61. Комментарии:Пока комментариев нет. Станьте первым! |